The testis-specific proteome
|
The main functions of the testis are the production of sperm (spermatogenesis), which is essential for reproduction, and synthesis of hormones that are important for the development of male sex characteristics. The vast majority of cells in the testis reside in the seminiferous duct, where spermatogenesis occurs. Additional testis-specific cell types are the hormone producing Leydig cells that are located outside the seminiferous ducts. The transcriptome analysis shows that 82% of all human proteins (n=19628) are expressed in the testis and 2200 of these genes show an elevated expression in testis compared to other tissue types.
An analysis of the genes with elevated expression in the testis shows that most of the corresponding proteins are involved in spermatogenesis and are thus expressed in the seminiferous ducts. In contrast, only a few of the elevated proteins in testis are expressed in the hormone producing Leydig cells.
|
- 1035 testis enriched genes
- Most of the enriched genes encode proteins involved in the spermatogenesis
- 2200 genes defined as elevated in the testis
- Most group enriched genes share expression with the fallopian tube
|
Figure 1. The distribution of all genes across the five categories based on transcript abundance in testis as well as in all other tissues.
2200 genes show elevated expression in the testis compared to other tissues. The three categories of genes with elevated expression in testis compared to other organs are shown in Table 1.
Table 1. Number of genes in the subdivided categories of elevated expression in testis
Category |
Number of genes |
Description |
| Tissue enriched |
1035 |
At least five-fold higher mRNA levels in a particular tissue as compared to all other tissues |
| Group enriched |
363 |
At least five-fold higher mRNA levels in a group of 2-7 tissues |
| Tissue enhanced |
802 |
At least five-fold higher mRNA levels in a particular tissue as compared to average levels in all tissues |
| Total |
2200 |
Total number of elevated genes in testis |
The list of tissue enriched genes (n=1035) includes previously characterized genes with cellular location and functions well in-line with the function of the testis, as well as a large number of genes with unknown function and expression pattern.
Table 2. The 12 genes with the highest level of enriched expression in testis. "Predicted localization" shows the classification of each gene into three main classes: Secreted, Membrane, and Intracellular, where the latter consists of genes without any predicted membrane and secreted features. "mRNA (tissue)" shows the transcript level as�TPM values,�TS-score (Tissue Specificity score) corresponds to the score calculated as the fold change to the second highest tissue.
Gene |
Description |
Predicted localization |
mRNA (tissue) |
TS-score |
| PRM1 |
protamine 1 |
Intracellular |
9152.7 |
5044 |
| PRM2 |
protamine 2 |
Intracellular |
7760.1 |
5039 |
| BOD1L2 |
biorientation of chromosomes in cell division 1-like 2 |
Intracellular |
423.7 |
4237 |
| AKAP4 |
A kinase (PRKA) anchor protein 4 |
Intracellular |
278.0 |
2780 |
| TPD52L3 |
tumor protein D52-like 3 |
Intracellular |
273.5 |
2735 |
| LELP1 |
late cornified envelope-like proline-rich 1 |
Intracellular |
779.6 |
2723 |
| TNP1 |
transition protein 1 (during histone to protamine replacement) |
Intracellular |
4375.5 |
2182 |
| TUBA3C |
tubulin, alpha 3c |
Intracellular |
513.4 |
2145 |
| ADAD1 |
adenosine deaminase domain containing 1 (testis-specific) |
Intracellular |
210.7 |
2108 |
| ACTL7A |
actin-like 7A |
Intracellular |
193.2 |
1933 |
| GK2 |
glycerol kinase 2 |
Membrane |
151.7 |
1517 |
| SPATA8 |
spermatogenesis associated 8 |
Intracellular |
402.0 |
1465 |
Some of the proteins predicted to be membrane-spanning are intracellular, e.g. in the Golgi or mitochondrial membranes, and some of the proteins predicted to be secreted can potentially be retained in a compartment belonging to the secretory pathway, such as the ER, or remain attached to the outer face of the cell membrane by a GPI anchor.
The testis transcriptome
An analysis of the expression levels of each gene makes it possible to calculate the relative mRNA pool for each of the categories. The analysis shows that 76% of the mRNA molecules in the testis correspond to housekeeping genes and that only 16% of the mRNA pool corresponds to genes categorized to be testis enriched, group enriched, or testis enhanced. Thus, most of the transcriptional activity in the testis relates to proteins with presumed housekeeping functions as they are found in all tissues and cells analyzed.
Protein expression of genes elevated in testis
In-depth analysis of the highly enriched genes in testis using antibody-based protein profiling allowed us to create a map of where
these proteins are expressed with regards to the various cell types in seminiferous ducts corresponding to the different phases of the
spermatogenesis including spermatogonia, spermatocytes, spermatids and mature sperms as well as Sertoli cells and Leydig cells.
Proteins specifically expressed in spermatogonia
Spermatogonia are diploid cells that form the basal layer of the seminiferous duct and present the initial phase of the spermatogenesis. The spermatogonia can be divided into two subtypes, type A cells that have stem cell like properties and maintain the spermatogonia population, and type B cells that produce primary spermatocytes.
Examples of genes expressed in spermatogonia include DMRT1, a transcription factor primarily expressed in the nuclei of spermatogonia that plays a key role in male sex determination and differentiation by controlling testis development and male germ cell proliferation, and the two genes PAGE1 and PASD1, two less well characterized proteins that belong to the group of cancer testis antigens.
Proteins specifically expressed in spermatocytes
Spermatocytes derive from type B spermatogonia and can be subdivided into primary spermatocytes that enter the first meiosis, and
secondary spermatocytes that enter the second meiosis to produce haploid spermatids. Several of the testis specific proteins localized to
spermatocytes are involved in testicular differentiation, proliferation and meiosis. Examples of genes specifically expressed in
spermatocytes include DAZL, a well-known RNA-binding protein that is essential for gametogenesis in
both males and females, and two poorly characterized genes TEX101 and
SHCBP1L.
Proteins specifically expressed in spermatids
Spermatids are derived from secondary spermatocytes and these cells are subdivided into early, round spermatids that are transcriptionally
active and late, elongated spermatids that are transcriptionally inert. The highly testis enriched proteins with known function and
predominant expression in spermatids are for example involved in conversion of nucleosomal chromatin and sperm development and
maturation. Examples of genes specifically expressed in spermatids include TNP1, which is involved
in the conversion of nucleosomal chromatin to the compact, non-nucleosomal form found in the sperm nucleus and
ACTL7B, a gene with only evidence of existence at the transcript level which encodes a putative
protein of unknown function. ACTL7B is suggested to be a member of a family of actin-related proteins. The formation of the acrosome, a
testis specific organelle required for sperm-egg interactions, is formed during the meiotic development of the late spermatid into mature
spermatozoa. An example of a gene specifically expressed in the acrosome is ACRV1. This gene is
expressed in the acrosomal vesicle during spermatogenesis and is associated with the acrosomal membranes and matrix of mature sperm.
The ACRV1 protein may be involved in sperm-zona binding or penetration.
Proteins specifically expressed in sperm
Following the maturation of late stage spermatids, the testicular end stage of spermatogenesis results in the formation of sperm. Sperm consists of uniflagellar cells that are localized along the lumen of seminiferous ducts. Specific proteins localized to mature sperm are involved in functions including sperm motility and male fertility. Examples of genes specifically expressed in sperm include PRM2, a protamine whose function is to substitute for histones in sperm and package sperm DNA into a highly condensed, stable and inactive complex, GAPDHS, an enzyme that catalyzes an important energy-yielding step in carbohydrate metabolism during spermatogenesis and AKAP4, suggested to be a major structural component of sperm fibrous sheath, which also plays a role in sperm motility.
Proteins specifically expressed in Sertoli and Leydig cells
The interaction between germinal cells and Sertoli cells is essential both for the development of testis and for the progress of the
spermatogenesis. Of the highly enriched genes in testis, only a few are specifically localized to Sertoli or Leydig cells, as compared to
the germ cell specific proteins. One example of a gene expressed in Sertoli cells is SLCO6A1,
currently with evidence only at transcript level. The INSL3 gene, specifically expressed in Leydig
cells, is suggested to be involved in the development of urogenital tract and involved in intra-abdominal testicular descent.
Genes shared between testis and other tissues
There are 363 group enriched genes expressed in the testis. Group enriched genes are defined as genes showing a 5-fold higher average level of mRNA expression in a group of 2-7 tissues, including testis, compared to all other tissues.
In order to illustrate the relation of testis tissue to other tissue types, a network plot was generated, displaying the number of commonly expressed genes between different tissue types.
Figure 2. An interactive network plot of the�testis�enriched and group enriched genes connected to their respective enriched tissues (grey circles).�Red�nodes represent the number of�testis enriched genes and�orange�nodes represent the number of genes that are group enriched. The sizes of the red and orange nodes are related to the number of genes displayed within the node. Each node is clickable and results in a list of all enriched genes connected to the highlighted edges. The network is limited to group enriched genes in combinations of up to�2�tissues, but the resulting lists show the complete set of group enriched genes in the particular tissue.
The network plot reveals that most group-enriched genes are shared with the fallopian tube (n=108) and the tissue that testis shares second most genes with is the cerebral cortex (n=45).
A Gene ontology-based analysis of the shared genes between testis and fallopian tube shows enrichment for genes related to cilia function and movement. One of the group enriched genes between testis and fallopian tube that is involved in cilia function and movement is DNAI1. This gene encodes a member of the dynein intermediate chain family and the encoded protein is part of the dynein complex in respiratory cilia.
For the shared genes between testis and cerebral cortex, there is no clear pattern of enriched GO terms.
SH3GL3 is an example of a group enriched gene in testis and cerebral cortex, and this gene is implicated
in endocytosis.
Testis function
The main functions of the testis are spermatogenesis, which is production of haploid germ cells essential for reproduction, and
synthesis of androgens (mainly testosterone) necessary for e.g. development of male sex characteristics. These two functions occur in the
two histologically different regions of the testis. Spermatogenesis takes place in seminiferous ducts, whereas the production of androgens
occurs in Leydig cells that are interspersed between the seminiferous ducts. During spermatogenesis, cells of seminiferous ducts undergo
meiosis which contains several specific events such as reduction of the number of chromosomes, condensation of nucleus and removal of
excess cytoplasm within the sperm. The sperm cells acquire unique morphological features and structures such as flagellum and acrosome that
are necessary for sperm motility and fertilization of the egg.
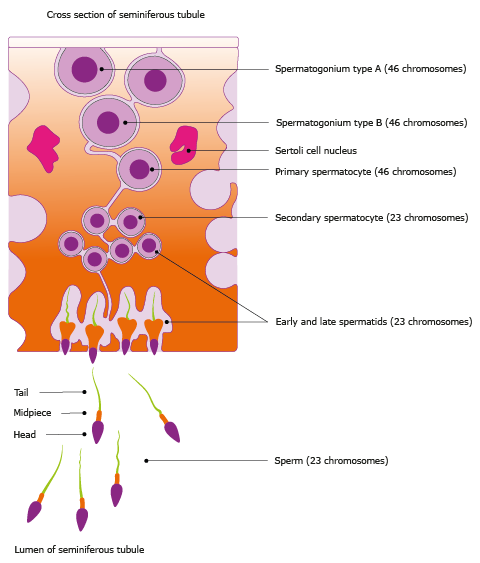
Figure 3. A schematic figure representing the cross section of a seminiferous tubule illustrating the stepwise
process of spermatogenesis. Initially, immature cells (spermatogonia) located close to the basal membrane divide by several rounds of
mitosis into primary and secondary spermatocytes that divide by meiosis to form spermatids. The final step of spermatogenesis takes place
in the lumen of the seminiferous tubule where spermatids differentiate into haploid sperm cells.
Testis histology
The outermost part of the testis is surrounded by the tunica albuginea which is a fibrous layer of connective tissue. The
testis is structured into lobules separated by fibrous septa where each lobule contains 1-4 seminiferous ducts. The seminiferous
ducts make up the majority of the testicular tissue and each seminiferous duct consists of germ cells and Sertoli cells. Germ c
ells undergo mitosis and meiosis and mature into sperm in the process of spermatogenesis. Spermatogenesis involves several complex
steps that can be morphologically determined and visualized in cross sections from seminiferous ducts. The different stages of
spermatogenesis include undifferentiated spermatogonia in the basal compartment followed by more luminal mature spermatocytes and
spermatids and the end product of this process which is mature sperm. The Sertoli cells are hormone producing non-dividing
columnar cells that are attached to the basement membrane and exhibit cytoplasmic extensions around the germinal cells. Sertoli
cells have irregular nuclei with a typical prominent nucleolus. Interspread between the seminiferous ducts are the
hormone-producing Leydig cells together with blood vessels, lymphatics, nerves and inflammatory cells.
The histology of human testis including detailed images and information about the different cell types can be viewed in the Protein Atlas Histology Dictionary.
Background
Here, the protein-coding genes expressed in the testis are described and characterized, together with examples of immunohistochemically stained tissue sections that visualize protein expression patterns of proteins that correspond to genes with elevated expression in the testis.
Transcript profiling and RNA-data analyses based on normal human tissues have been described previously (Fagerberg et al., 2013). Analyses of mRNA expression including over 99% of all human protein-coding genes was performed using deep RNA sequencing of 172 individual samples corresponding to 37 different human normal tissue types. RNA sequencing results of 10 fresh frozen tissues representing normal testis was compared to 162 other tissue samples corresponding to 36 tissue types, in order to determine genes with elevated expression in testis. A tissue-specific score, defined as the ratio between mRNA levels in testis compared to the mRNA levels in all other tissues, was used to divide the genes into different categories of expression.
These categories include: genes with elevated expression in testis, genes expressed in all tissues, genes with a mixed expression pattern, genes not expressed in testis, and genes not expressed in any tissue. Genes with elevated expression in testis were further sub-categorized as i) genes with enriched expression in testis, ii) genes with group enriched expression including testis and iii) genes with enhanced expression in testis.
Human tissue samples used for protein and mRNA expression analyses were collected and handled in accordance with Swedish laws and regulation and obtained from the Department of Pathology, Uppsala University Hospital, Uppsala, Sweden as part of the sample collection governed by the Uppsala Biobank. All human tissue samples used in the present study were anonymized in accordance with approval and advisory report from the Uppsala Ethical Review Board.
Relevant links and publications
Uhlén M et al, 2015. Tissue-based map of the human proteome. Science
PubMed: 25613900 DOI: 10.1126/science.1260419 Yu NY et al, 2015. Complementing tissue characterization by integrating transcriptome profiling from the Human Protein Atlas and from the FANTOM5 consortium. Nucleic Acids Res.
PubMed: 26117540 DOI: 10.1093/nar/gkv608 Fagerberg L et al, 2014. Analysis of the human tissue-specific expression by genome-wide integration of transcriptomics and antibody-based proteomics. Mol Cell Proteomics.
PubMed: 24309898 DOI: 10.1074/mcp.M113.035600 Djureinovic D et al, 2014. The human testis-specific proteome defined by transcriptomics and antibody-based profiling. Mol Hum Reprod.
PubMed: 24598113 DOI: 10.1093/molehr/gau018
Histology dictionary - testis
|